1オートファジーって何?
オートファジー=Autophagyのautoはギリシャ語で「自己」、phagyはphage等と同類で「食べる」の意です。日本語では、自食作用とか自己貪食などというおどろおどろしい訳語が使われますが、要するに細胞が自己成分を分解する機能のことです。近年オートファジーの研究は飛躍的に発展し、オートファジーが発がん、神経変性疾患、2型糖尿病等の生活習慣病、心不全、腎症、感染症、各種の炎症など、さまざまな重要疾患の発症を抑止していること、また発生・分化、老化、免疫などにおいて重要な生理機能を持つことが明らかになり、オートファジー研究は現在大きな注目を集めている研究分野です。
細胞を構成するタンパク質などの高分子は、時間がたつと何となく壊れるのではなく、一定期間後細胞によって能動的に分解されており、合成と分解のバランスによって生命は成立しています。オートファジーは、プロテアソーム系と並ぶ主要な細胞内分解システムです。前者がいわゆる短寿命タンパク質の分解を司るのに対し、後者は細胞構成成分の多数を占める長寿命タンパク質の分解を遂行します。オートファジー(注1)の実態は、細胞質中のタンパク質等をリソソームに運ぶメンブレントラフィックで、それもメンブレントラフィック入門で述べた小胞輸送とは異なる極めてユニークな方式のものです(図1)。
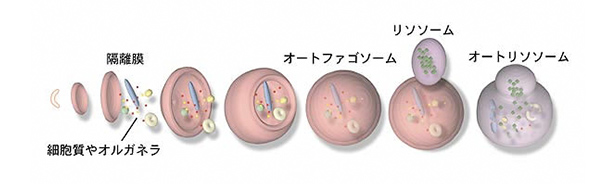
- 図1:オートファジーの膜動態の模式図
- 隔離膜が細胞質やオルガネラの一部を取り囲み、オートファゴソームとなります。そこにリソソームが融合し、オートリソソームが形成されると取り込まれた内容物が分解されます。
まず細胞質に隔離膜と呼ばれる扁平な膜区画が出現、湾曲しながら伸長し、細胞質や場合によってはミトコンドリアやペルキシソーム等のオルガネラを包み込みます。最後に隔離膜の末端同士が融合し直径約1μmの閉じた2重膜構造が完成します。それがオートファゴソームです。続いてその外膜に消化オルガネラであるリソソームが融合しオートリソソームになると、流入した加水分解酵素が内膜と包み込んだ内容物を消化します。これらの過程は、数十分で進行します。オートファゴソームはれっきとしたオルガネラですが、常に存在するミトコンドリアやゴルジ体などの他のオルガネラと異なり、必要に応じて作られては消える大変ユニークなオルガネラです。プロテアソームが、ユビキチン化された標的タンパク質を一分子ずつ選択的に分解するのに対し、オートファジーでは細胞内のある空間がごっそりリソソーム酵素で分解されるためバルク(要するに大雑把な)分解系と呼ばれます。
オートファジー研究の歴史は古く、1950年代には哺乳動物細胞において電子顕微鏡下で観察されていました。ところが分子レベルでの解析は大きく遅れ、つい最近までほとんどブラックボックスと言っていい状態でした。分解機構としてもメンブレントラフィックとしても謎だらけの存在に一筋の光が当たったのは、現・東京工業大学の大隅良典特任教授らが1993年に出芽酵母のオートファジー不能変異株群・atgの同定に成功したときでした。このブレイクスルーは、その後の怒濤のような研究展開のきっかけとなりました。酵母Atgのホモログ解析から、哺乳動物オートファジーの解析も進み、酵母と哺乳動物でオートファジーの分子機構がよく保存されていること、また、哺乳動物においてオートファジーが様々な生理機能を持つことが明らかになってきました。
注1:オートファジーには、シャペロン介在性オートファジー、ミクロオートファジー、マクロオートファジーの3つの形式が知られています。そのうちマクロオートファジーが最も主要で単にオートファジーと呼ばれる場合が多いので、ここでもオートファジーはマクロオートファジーを指すものとします。
2哺乳動物オートファジーの分子機構
私達はAtgの哺乳動物ホモログの解析にいち早く取り組み、オートファジーの分子基盤を探ってきました。以下にその成果の概略を述べます。まず私達は、酵母Atg8のホモログLC3がオートファゴソーム膜に特異的に結合することを見いだしました(EMBO J, 19, 5720-8, 2000)。LC3は細胞質で合成後、直ちにC末端22残基がシステインプロテアーゼであるAtg4により切断除去されます。その結果生じたLC3-Iは、細胞質に拡散しています。LC3-Iが、リン脂質分子の1つフォスファチジルエタノールアミン(PE)に共有結合するとLC3-IIとなり、隔離膜/オートファゴソーム膜に繋ぎ止められます(図2)。LC3-IIの量はオートファゴソーム形成と正の相関を示すため、オートファジーの誘導や抑制を評価するための実験手法としてLC3-II量の測定が世界中で使われています。また、LC3は信頼できる哺乳動物のオートファゴソームマーカーとして顕微鏡観察によく用いられています。そのお陰で、私達のLC3の論文は引用が2,900を超え(2014年の時点で)、オートファジー分野では世界2位になっています。
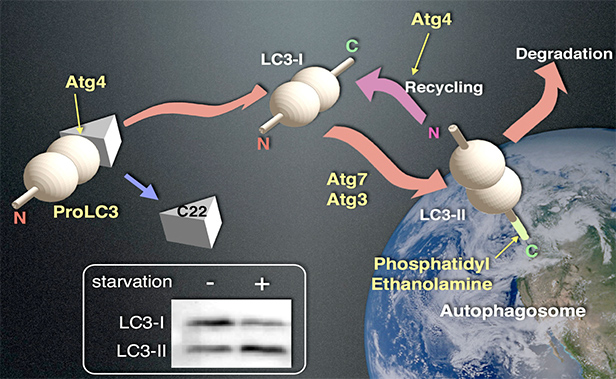
- 図2:LC3のオートファゴソーム膜への結合
- LC3はProLC3(左)として翻訳された後、直ちにAtg4によってC末端22アミノ酸が除去され、LC3-I(中)となります。LC3-IはまずAtg7に、続いてAtg3に受け渡された後、隔離膜/オートファゴソーム膜のリン脂質分子フォスファチジルエタノールアミン(Phosphatidylethanolamine (PE))にC末端で共有結合してLC3-II(右)となります。オートファゴソームの外側のLC3-IIはAtg4によってPEから切断され、再びLC3-Iに戻ることが出来ますが、オートファゴソームの内側のLC3-IIは、オートファゴソームがリソソームと融合すると分解されます。
左下図:LC3-IとLC3-IIは、ウエスタンブロット法を用いると分離して検出することができます。LC3-IIの量はオートファゴソームの数と相関するため、オートファジー活性の指標として使われます。
私達はまた、酵素活性を持たない変異型Atg4を細胞に過剰に発現させるとLC3-IIの生成が阻害されることを見出し、これを実験ツールとして利用することで、LC3が隔離膜閉鎖に働くことを明らかにしました(Mol Biol Cell, 19, 4651-59, 2008)。つづいて、LC3が隔離膜/オートファゴソーム膜に共有結合するという上記の反応が、Atg16L1複合体(Atg16L1, Atg5, Atg12からなる複合体)によって促進されることを示しました(Mol Biol Cell, 19, 2092-100, 2008; Nature, 456, 264-68, 2008)。Atg16L1複合体は隔離膜に結合するという性質を持っていますが、Atg16L1複合体を人為的に細胞膜に繋ぎ止めると、細胞膜でLC3-II化が起こりました。このことから、LC3自身に特定の膜を選ぶ能力があるのではなく、Atg16L1が隔離膜を認識してそこでLC3-II化を起こさせると考えられます。
オートファゴソーム膜にはフォスファチジルイノシトール3リン酸(PI3P)というリン脂質が含まれ、それを産生するⅢ型PI3キナーゼ(PI3K)という酵素がオートファゴソーム形成に必須であることが分かっています。この酵素はBeclin1(Atg6の哺乳類ホモログ)、Vps15と複合体を形成することを私達は見つけました(EMBO rep, 2, 330-335, 2001)。さらに私達は、Beclin1と相互作用する新規タンパク質Atg14LとRubiconを同定し、前者がオートファゴソーム形成を促進する一方、後者はオートファゴソームとリソソームの融合を抑制するという興味深い制御の存在を示しました。また、Rubiconを含む複合体はエンドサイトーシス経路における輸送も抑制します。つまり、Beclin1-Vps15-PI3K複合体は、結合する相手を変えることによりオートファジーだけでなくエンドサイトーシス経路も制御することが分かりました(Nat Cell Biol, 11, 385-96, 2009)。Beclin1は発がん抑制能を持つことが以前から知られていましたが、このような複雑なメンブレントラフィックの制御を通して発がん抑制に関与しているのかも知れません。
私達は、数十年に亘って論争されてきた「オートファゴソーム膜の起源」の謎についても解析を進めてきました。必要に応じて現れるオートファゴソームがどこでどのようにしてできているのかは、決定的な証拠が無く結論の出ない状態が続いていましたが、2008年にオートファゴソームが小胞体の近くで形成されることが、LC3やDFCP1(小胞体に局在するPI3P結合タンパク質)のライブ撮影により示されました。私達の解析結果からも、オートファゴソームの小胞体起源説を強く支持する結果が得られました。私達と長浜バイオ大学の山本章嗣教授との共同研究で、隔離膜の電子線トモグラフィー3D画像を構築したところ、隔離膜は2枚の小胞体シートに挟まれており、しかも隔離膜と小胞体が一部で連続しているという観察結果が得られました。この結果から、隔離膜が2枚の小胞体シートに挟まれて成長する可能性が示唆され、私達はこれを「小胞体揺りかごモデル」と名づけました(Nat Cell Biol. 11, 1433-7, 2009)(図3)。私達はまた、Atg14Lが小胞体に存在することを明らかにし、Atg14Lの小胞体局在がオートファゴソーム形成誘導に必要であることを示しました(J Cell Biol. 190, 511-21, 2010)。飢餓などのオートファジー誘導刺激が起こると、Atg14を含むPI3K複合体が小胞体上に点状に集合し、その結果、小胞体膜のその箇所でPI3Pの局所濃度が上昇し、小胞体膜が変形し隔離膜が形成されると考えられます。その後、小胞体起源説に対しミトコンドリア起源説が提唱され対立が起こりましたが、私達のさらなる研究により、隔離膜が形成される小胞体上の箇所はミトコンドリアと小胞体の接触部位であることが示され、小胞体起源説とミトコンドリア起源説の対立に終止符が打たれました(Nature 495, 389-393, 2013)。この研究においては、小胞体とミトコンドリアが接している部位でオートファゴソームが作られていることを、世界でも類を見ない蛍光3色同時ライブ撮影などを使って明らかにし、大きな話題となりテレビニュースでも取り上げられました(図4)。この結果は、2つの説の両方が正しいことを示しただけでは無く、小胞体とミトコンドリアという機能も由来も全く異なる2つの独立したオルガネラが、協働して第3のオルガネラ・オートファゴソームを作ると言う、従来のオルガネラ観を覆す新しいパラダイムを提示した点で細胞生物学上の大きな発見だと言えます。
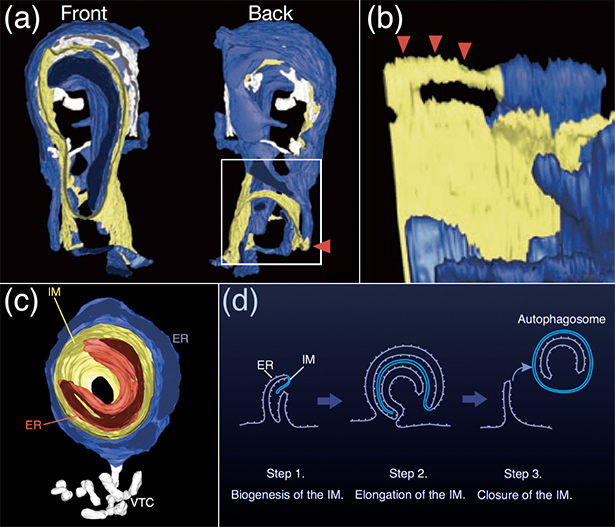
- 図3:小胞体におけるオートファゴソーム形成
- (a, b): 電子線トモグラフィー法により構築された、小胞体(青色)と隔離膜(黄色)の3D画像。隔離膜と小胞体がつながっている様子が赤色矢印で示されています。(b)は(a)の四角部分を拡大し、異なる角度から見た図です。スケールバー=100 nm。©:この3D画像では、隔離膜(黄色)が2つの小胞体シート(青色と赤色)に挟まれているのが示されています。近くには、小胞体に特徴的な構造であるVesicular tubular clusters (VTC)(白色)と呼ばれる構造が見られます。(d): 小胞体揺りかごモデル。隔離膜(IM)が小胞体(ER)の一部から伸長し(Step 1)、2枚の小胞体シートに支えられて成長し(Step 2)、最後に閉じてオートファゴソームとなる(Step 3)というモデルを提唱しました。(Originally published in Nature Cell Biology, 11, 1433-7, 2009,doi:10.1038/ncb1991)
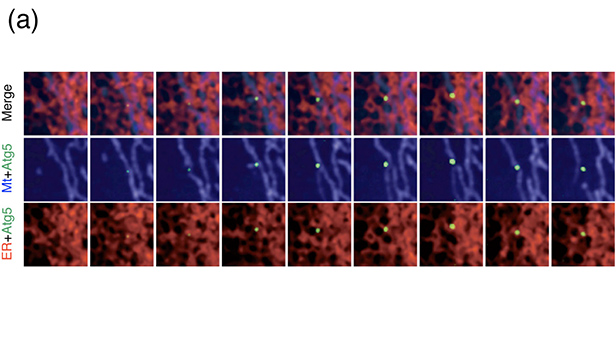
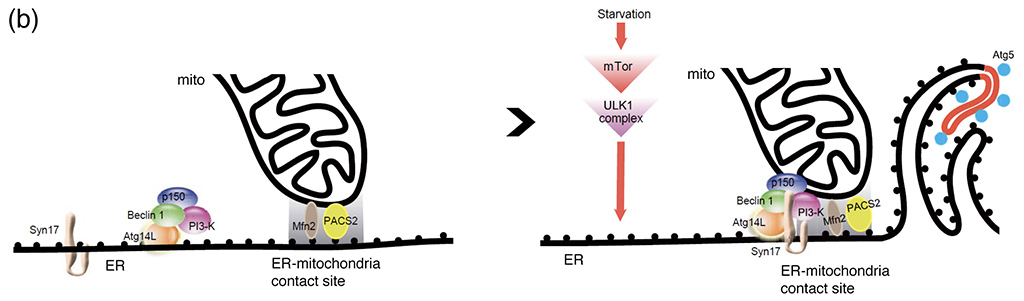
- 図4:小胞体-ミトコンドリアコンタクトサイトにおけるオートファゴソーム形成
- (a):小胞体(赤色)とミトコンドリア(青色)が接している場所から、隔離膜マーカーのAtg5(緑色)が現れることが示されました。めまぐるしく動くこれらのオルガネラの位置関係を正確に捕らえるためには、3台の高感度CCDカメラを用いた同時3色蛍光観察が必要でした。(b):栄養飢餓などの刺激により、mTorと ULK1複合体を介してオートファジーが誘導されます。このときSyntaxin17と呼ばれるタンパク質がAtg14L-PI3K複合体を小胞体とミトコンドリアのコンタクトサイトに連れてくることが分かりました。PI3KはそこでPI3Pを産生し、隔離膜の形成が始まると考えられます。この研究結果はNHKのニュースでも放送されました。
(Originally published in Nature, 495, 389-393, 2013 doi:10.1038/nature11910)
以上のように、私達はオートファゴソーム形成の分子機構の一端やオートファゴソーム膜の起源を明らかにしてきました。現在さらに、このユニークなメンブレントラフィックの分子機構の解明を目指し研究を続けています。
3哺乳動物オートファジーの生理機能と疾患との関係
オートファジーの研究は近年急速に進み、その結果、オートファジーが生体にとって極めて重要で多岐に亘る機能を持つことが判ってきました(図5)。オートファジーの役割として当初から考えられていたのは、飢餓に陥ったときの栄養源確保です。細胞は飢餓やその他のストレスに曝されると、自己の一部をオートファジーによって分解し栄養源にすることでサバイバルをはかります。一方で、最近の研究で明らかになってきたオートファジーの重要な役割として、細胞の新陳代謝(代謝回転)と細胞にとって不要あるいは有害な物質の除去があります。ヒトなどの動物は食物としてたんぱく質を摂取・分解しますが、それとは別に自己の体たんぱく質を一日に1-2%(成人男性で約200g)も分解しており、しかもそのうちの70-80%は再びたんぱく質合成に使われます。つまり、毎日壊しては造るということを繰り返しているのです。何故生物はこのような一見無駄なことをしているのでしょう? その答えは、細胞の主要な分解系であるオートファジーの研究から明らかになりました。オートファジーによるこの新陳代謝が停止すると、細胞が弱ったり死んだりして様々な病気〜発がん、神経変性疾患、2型糖尿病等の生活習慣病、心不全、腎症、感染症、各種の炎症等々〜になってしまうのです。オートファジーはこのような日々の新陳代謝に加えて、細胞にとって危険な傷のついたミトコンドリア、変性疾患の原因となる凝集しやすいタンパク質、細胞に侵入した病原体などが現れると積極的に除去することでやはり上記のような疾患や感染症を防ぎます。日常的なオートファジー(基底オートファジーと呼ばれます)や飢餓時のオートファジーは非選択的な過程で細胞質にあるものをほぼランダムに分解しますが、有害物の除去は選択的な狙い撃ちの過程であることが判ってきました。その他にも、オートファジーは幹細胞が分化するときの古いオルガネラ除去による細胞内体制転換、細胞質抗原のクラスII MHCによる細胞表面での提示、細胞質の物質の細胞外への分泌などにも働いています。このようにオートファジーは、細胞の生存や恒常性維持に欠かせない守護神のような存在であり、日々生命を支え様々な病気から我々を守ってくれているのです。オートファジーが関わる疾患は社会的に重要なものが多く、そのために大きな注目を集めるようになりました。
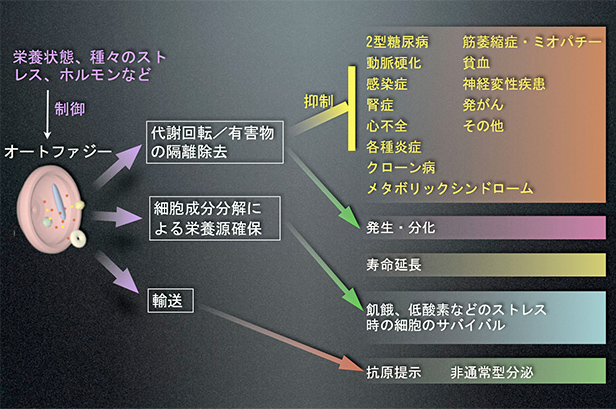
- 図5:オートファジーの機能
- オートファジーの主な働きとして、飢餓時の栄養源確保と、常に少しずつ起こっている代謝回転、細胞内に現れた有害物の隔離除去などがあります。特に、代謝回転と有害物の隔離除去は、生活習慣病を含む数多くの疾患発症を抑制していることが最近の研究で明らかになり、注目を集めています。
私達はオートファジーの機能的意義の究明に向けて、オートファジーが持つ有害物質の除去という役割を、特に疾患との関連に着目し研究してきました。私達はまず、細胞質に侵入した病原性細菌を巨大なオートファゴソームが捕獲し殺すことを世界で初めて見つけました(Science, 306, 1037-40, 2004)(図6)。この発見はオートファジーが自食という定義を越えて、非自己である病原体を攻撃し自然免疫としても働くことを示す画期的な成果でした。この発見以降、様々な細菌、ウイルス、原生動物などの病原体についてオートファジーとの関係が次々報告されています。
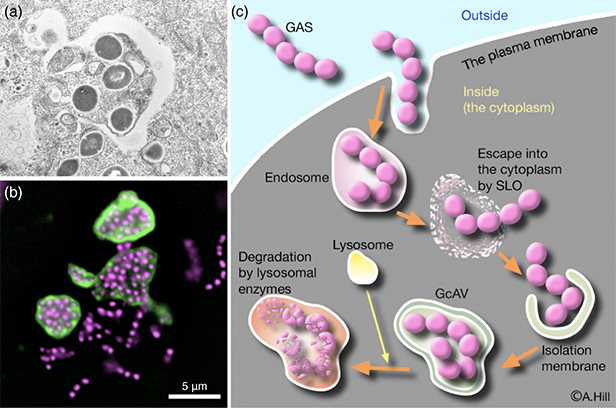
- 図6:オートファジーによるA群レンサ球菌Group A Streptococcus (GAS)の殺菌
- (a):電子顕微鏡で示された、GASの菌体が2重膜によって包まれている様子。
(b):オートファゴソームマーカーのLC3(緑)を用いた蛍光観察によって、GAS(マゼンタ)がオートファゴソームに包まれていることが示されました。
(Originally published in Science, 306, 1037-40, 2004 DOI: 10.1126/science.1103966)
(c):細胞内に侵入してきたGASは、細菌毒素ストレプトリシンO(SLO)を用いてエンドソーム膜を壊し、細胞質に逃げ出そうとしますが、そのとき巨大なオートファゴソーム(GAS containing LC3-positive autophagosome-like vacuoles (GcAVs))がGASを捕まえ、続いてリソソームと融合することでGASを殺菌します。
一般にオートファゴソームはランダムに細胞質中のものを取り込むと考えられていましたが、病原体などの有害物を捕獲するときには、オートファゴソームは明らかに狙い撃ちしていました。私達は、オートファジーが病原体を狙い撃ちする仕組みについても研究を進めてきました(PLoS Pathog. 5, e1000670, 2009; Mol Biol Cell. 22, 2290-300, 2011; J Cell Biol, 203, 115-28, 2013)。これらの研究の結果まず判ったのは、細胞外からエンドサイトーシス経路を通して侵入してきた細菌が、エンドソーム膜を破って細胞質に逃げようとしたり、物質を送り込むために穴を開けたりした時に、オートファゴソームが選択的に形成されるということです。この事実から私達は、もしかしたら細胞は菌を認識しているのでは無く、損傷したエンドソーム膜を標的にしているのかもしれないと考えました。それを実証するために、直径数マイクロメートルのポリスチレンでできた人工ビーズにトランスフェクション試薬をまぶして細胞に取り込ませました。このようなビーズがオートファジーの標的になることが既に報告されていて、それはビーズが入り込んだエンドソームの膜を破るためではないかと考えたのです。予想は的中し、ビーズを含むエンドソーム膜が破れて、オートファジー開始に必要なAtgタンパク質が破れたエンドソーム膜に集合してくる様子が観察されました(図7)。このとき、破れたエンドソーム膜上のタンパク質にはユビキチン化と呼ばれるタンパク質修飾が起きていました。ユビキチン化は、上述のプロテアソーム系において分解されるタンパク質の標識として使われていることで有名ですが、近年の研究で、オートファジーによる分解標識としても使われていることが明らかになっています。しかし、ユビキチン化がどのようにしてオートファジーを開始させるのかは多くが謎に包まれていました。私達は、オートファジー開始に必要なAtgタンパク質が、破れたエンドソーム膜に起こったユビキチン化を認識して集合することを示し(Atg16L1についてはユビキチンとの結合という直接的なメカニズムの存在も明らかになりました)、ユビキチン化がオートファジーを開始させる分子機構の一端を解明しました。私達が見つけたオートファジーによる細菌狙い撃ちの仕組みは、NHKの「サイエンスZERO」で放映されました。
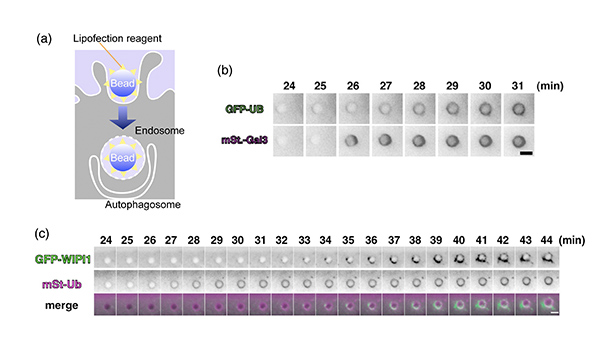
- 図7:ポリスチレンビーズを用いたオートファゴソーム形成誘導実験
- (a):トランスフェクション試薬をまぶしたビーズを細胞に投与すると、エンドサイトーシスによりビーズが細胞内に取り込まれます。ビーズを含むエンドソームが破れた時にオートファゴソーム形成が起こります。
(b):実際に細胞内に取り込まれたビーズをライブ撮影してみたところ、ビーズを含むエンドソーム膜が破れ(下段:mSt-Gal3は、エンドソーム膜が破れた時に集まる分子です)、続いてユビキチン化(上段:GFP-UB)が起こりました。ユビキチン化の後にオートファゴソームマーカーのLC3が集まって来ることも確認しています。
(c):ビーズを含むエンドソーム膜が破れた時にユビキチン化(マゼンタ)が起こり、続いて各種のAtgタンパク質(ここではAtg18の哺乳類ホモログであるWIPI1(緑))が集まってくるところが捉えられました。スケールバー=3μm。
(Originally published in Journal of Cell Biology, 203, 115-28, 2013, doi: 10.1083/jcb.201304188)
以上のように私達は、細胞は実は菌に気付いておらず破れたエンドソームを何とかしようとしてオートファジーを起こしているのかも知れない、という興味深い結論に辿り着きました。もちろん私達の結果は、細菌自身がオートファジーの標的になる可能性を否定するものではありませんが、何故オートファジーは様々な病原体を攻撃できるのか(例えば表面の分子構成が全く異なるグラム陽性菌と陰性菌の両方が標的になります)という疑問はこれで解けたのかも知れません。私達は、損傷エンドソームがオートファジーの標的になると言う事実から、損傷リソソームについても解析を行いました。実はリソソームは生理的にしばしば損傷を受けます。尿酸結晶、シリカ、コレステロール結晶、アミロイドたんぱく質など様々な因子がエンドサイトーシスで細胞に取り込まれ、最終到達点であるリソソームを傷つけます。私達は、エンドソームの場合同様に傷ついたリソソームにユビキチンが結合し、選択的にオートファゴソームがそれを除去することを見出しました(EMBO J, 32, 2336-47, 2013)(図8)。さらに高尿酸血症にしたマウスの尿細管のオートファジーを止めると、尿酸結晶によって破れたリソソームが除去されず腎症が悪化することも阪大腎臓内科との共同研究で明らかにしました。尿酸結晶、コレステロール結晶、ヒト膵アミロイドペプチドなどは痛風、動脈硬化、2型糖尿病などの生活習慣病の原因因子ですが、リソソームに傷をつけます。そして生活習慣病になりやすくなる過栄養が、オートファジーを低下させることが判ってきました。生活習慣病の根底には、損傷リソソームを除去するオートファジー(リソファジーと呼んでいます)の低下による悪循環があるのかもしれません。
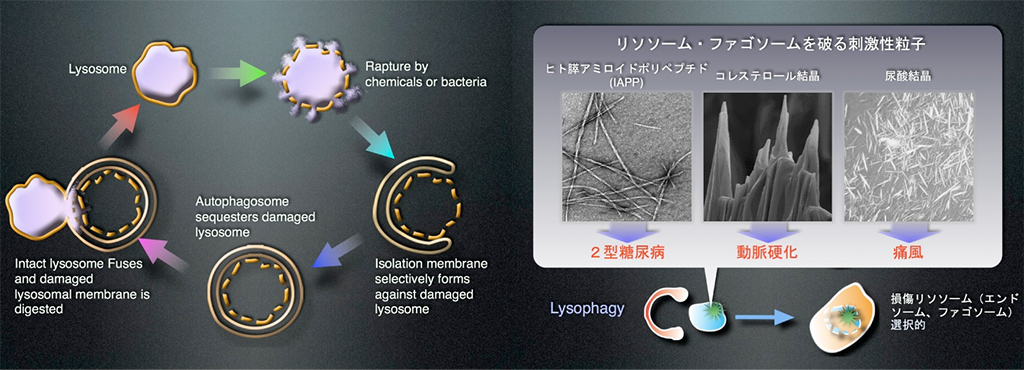
- 図8:リソファジーの模式図
- 左図:リソソームが損傷を受けると、ユビキチンとAtgタンパク質が損傷リソソームに集まり、オートファゴソーム形成が起こります。オートファゴソームが損傷リソソームを包みこんで隔離し、そこに別の(損傷を受けていない)リソソームが融合することで、損傷リソソームが除去されます。
右図:ヒト膵アミロイドポリペプチド(IAPP)、コレステロール結晶、尿酸結晶などの刺激性粒子は過栄養によって形成され、2型糖尿病、動脈硬化、痛風などの生活習慣病の原因となります。これらの刺激性粒子はリソソーム(エンドソーム、ファゴソーム)の膜損傷を引き起こしますが、このときにリソファジーが損傷リソソームを除去することで生活習慣病の発症や悪化を防いでいる可能性があります。
また私達は、オートファジーが変性疾患(アルツハイマー病や狂牛病などが有名)の原因となる凝集しやすい異常蛋白質を選択的に分解し、その蓄積・凝集を防ぐ働きもあることをいちはやく示しています(J Biol Chem, 281, 4467-76, 2006)。このように私達は、細胞にとって招かれざる客である病原体、損傷リソソーム、易凝集性たんぱく質などの有害物をオートファジーが選択的に除去し細胞を保護していることを示し、その分子機構の解明にも貢献してきました。なお損傷ミトコンドリアのオートファジーによる除去(ミトファジー)も知られているので、オートファジーは壊れてしまったオルガネラを処理する普遍的装置なのでしょう。
4今行っていること
急成長するオートファジーの分野で、当研究室はパイオニアとして人まねではない先端的な研究を展開しています。駆使する技法も、生化学、分子生物学、細胞工学など多面的で、特にイメージングに力を入れておりレーザー顕微鏡、超解像顕微鏡、電子顕微鏡を教室で保有し世界有数の設備と技術を誇っています。現在は分子機構や生理機能の解析を行うと同時に、臨床医学の各教室との共同研究を多数積極的に展開し、生活習慣病やがんなどの疾患における役割を精力的に研究しています。そして将来の創薬を目指し、オートファジーを制御する薬剤同定のためのスクリーニングを実施しています。
- 文責:吉森